
Paměť a její poruchy
:
F. Koukolík
:
Primář: MUDr. František Koukolík DrSc.
; Národní referenční laboratoř prionových chorob
; Fakultní Thomayerova nemocnice s poliklinikou, Praha
; Oddělení patologie a molekulární medicíny
:
Prakt. Lék. 2008; 88(2): 82-87
:
Various Specialization
Paměť v lidském mozku není jednotný systém. Rozšířená klasifikace rozlišuje krátkodobou (pracovní) paměť a vědomou, dlouhodobou, neboli deklarativní paměť pro fakta, to je sémantická paměť a paměť pro události, které se říká epizodická a autobiografická. Existuje skupina nevědomých pamětí, jejich představiteli jsou procedurální paměť, priming, pavlovovské podmiňování a emoční podmiňování. Amnézie je porucha deklarativní paměti. Anatomická klasifikace amnézií rozlišuje amnézii hipokampální, diencefalickou, dále amnézii při poškození bazálního telencefala a některých systémů bílé hmoty mozku. Praktický lékař se běžně setkává s vývojem amnézie při minimální kognitivní poruše a Alzheimerově nemoci.
Klíčová slova:
paměť krátkodobá, deklarativní, sémantická, epizodická, amnézie
Úvod
Paměť je naším jediným bohatstvím. Teprve přijdeme-li o ni, stanou se z nás skuteční žebráci“, napsal J.E.D. Esquirol (1772–1840), velký francouzský psychiatr, jeden ze zakladatelů vědy o stárnutí mozku a demencích.
Paměť není jednotná funkce. Jestliže mluvíme o krátkodobé, neboli pracovní paměti, dále o deklarativní, neboli vědomé dlouhodobé paměti a jejích podobách nazvaných epizodická a sémantická paměť, stejně jako mluvíme-li o skupině nevědomých pamětí, mluvíme zároveň o odlišných neuronálních systémech i s jejich úzkými profily informačního chodu (tab. 1). Úzké profily informačního chodu jsou místa jejichž poškození znamená poškození funkce celého systému. Příkladem úzkého profilu informačního chodu deklarativní paměti je hipokampus.

Paměťové „moduly“ lidského mozku
Existuje řada možných klasifikací paměti (5, 6, 9). Výhodou klasifikace uvedené v tab. 1 (12) je přehlednost a vazba na neuronální systémy.
Pracovní paměť je krátkodobá paměť uchovávající informace asi 60 s. S oblibou se přirovnává k tabuli, na kterou se něco napíše a v krátké době vymaže, aby bylo možné napsat něco jiného. Informace mohou z pracovní paměti vymizet nebo se uložit do dlouhodobé paměti.
Dlouhodobá paměť má vědomou a nevědomou podobu.
Vědomé dlouhodobé paměti se říká deklarativní. Ta se jednak týká událostí včetně autobiografických, to je epizodická paměť, lze ji přirovnat k palubnímu deníku, jednak fakt a souvislostí, tu je možné přirovnat k encyklopedii, název zní sémantická paměť.
Existuje skupina nevědomých pamětí.
Procedurální paměť, též habit, je paměť pro dovednosti. Zpočátku se vědomě namáhavě učíme, například chodit, psát, jezdit na bruslích, plavat, psát na stroji, řídit automobil, všechna možná řemesla. Po nějaké době vědomého učení se procesy automatizují, při jejich výkonu na ně nemyslíme. Touto pamětí se budeme zabývat v budoucím článku o hybnosti a lateralitě.
Priming, doslovně rozněcování, je nevědomá paměť, bez které by bylo užití naučeného materiálu trvalé namáhavé. Smyslem primingu je „energetická úspora“.
Dalším druhem nevědomé paměti je pavlovovské podmiňování, které vypadá na první pohled jednoduše, jeho neuronální mašinérie je však velmi složitá. Popíšeme ji ve článku o motorice. Klíčové pro strukturu osobnosti a citového života jsou emoční podmiňování a emoční paměť, která má nevědomou i vědomou složku. Budeme se jí věnovat v budoucích článcích o citovém a sociálním mozku.
Pracovní paměť
Kognitivní věda říká, že pracovní paměť má tři složky:
- fonologickou smyčku,
- vizuospaciální náčrtník, a
- centrální výkonnostní složku (1).
Fonologická smyčka je systém, v němž se ukládají zvukové neřečové a řečové informace, které se, pokud nejsou opakovány, ztrácejí během dvou až tří sekund. Lidé si mohou tyto informace nehlasným opakováním udržovat dlouhodobě. Kapacita pracovní paměti je omezená. Mnohoslabičná slova se udržují obtížněji než slova jednoslabičná. PET dokládá, že činnost fonologické smyčky souvisí s aktivací levostranné kůry v bezprostředním okolí Sylviovy rýhy. Nehlasné opakování souvisí s aktivací Brocovy oblasti (BA 44, BA 45 vlevo), kromě toho se aktivuje i levostranná premotorická (BA 6 na zevní ploše hemisféry) a doplňková motorická kůra (BA 6 na vnitřní ploše hemisféry, obr. 1).

Vizuospaciální náčrtník je architektonicky i funkčně složitější než fonologická smyčka. Zobrazovací metody ukazují, že jeho činnost souvisí s aktivací zrakové kůry týlních laloků i kůry temenních i čelních laloků.
Centrální výkonnostní složka je vázána na činnost předních a horních zevních částí prefrontální kůry. Čelní kůra aktivovaná při zátěži pracovní paměti se podílí na integraci korových smyslových informací přicházejících ze zadních a spodních částí mozku. Aktivita čelní kůry roste úměrně stupni zátěže pracovní paměti. Tento popis pracovní paměti však je učebnicové zjednodušení. Dokazují to experimenty zatěžující pracovní paměť zvané „n-back“ úloha (doslovně „n-krát nazpět“, přičemž n = 1, 2, 3…).
Neuronální systémy, jejichž činnost odpovídá na zátěž pracovní paměti, jsou velmi rozsáhlé a jejich „uzly“ neslouží výlučně úkolům zatěžujícím pracovní paměť. Kromě toho záleží na druhu zatěžující úlohy. Úlohy zatěžující zrakovou a prostorovou pracovní paměť aktivují poněkud jiné oblasti čelní kůry než úlohy zatěžující zvukovou a slovní pracovní paměť.
Epizodická paměť
Projevem činnosti epizodické paměti jsou například vzpomínky na vlastní promoci 10. 6. 1965 dopoledne. Epizodická paměť je závislá na činnosti neuronální sítě, jejímiž zúženými profily jsou hipokampální formace společně s přilehlou spánkovou kůrou a prefrontální kůra.
Kódování slovních informací do epizodické paměti aktivuje převážně levostrannou dolní prefrontální kůru. Kódování neverbálních informací do epizodické paměti aktivuje převážně pravostrannou dolní prefrontální kůru. Obsah kódované informace ovlivňuje i činnost mediálního částí spánkového laloku, zejména hipokampální formace:
- kódování slovní informace aktivuje převážně levostrannou,
- kódování zrakových scén a zrakově pozorovaných předmětů aktivuje mediální části spánkového laloku oboustranně,
- kódování tváři aktivuje především stranu pravou.
Relativní novost kódované informace moduluje činnost jak dolních prefrontálních, tak mediálních temporálních korových oblastí účastnících se kódování paměti. Aktivace těchto korových oblastí je větší, jestliže je kódovaná informace nová.
Experiment sledující kódování dvou druhů zrakových, tj. neslovních informací do epizodické paměti, a to domů a tváří, prokázal, že úspěšné zakódování dvojice tvář-dům odpovídá míře aktivity levé dorsolaterální prefrontální kůry (11).
Při vybavování dat ze slovní epizodické paměti se aktivuje zejména pravostranná prefrontální kůra. Přitom záleží na kontextu vybavované informace. Jestliže je zátěž při experimentu zaměřena na monitorování kontextu vybavované informace, roste aktivita střední části pravostranné dorsolaterální prefrontální kůry. Jestliže se vybavují informace, které nejsou závislé na zadání úlohy v tom smyslu, že je není nutné monitorovat, aktivuje se ventrálnější oblast pravostranné prefrontální kůry. Naproti tomu precuneus (BA 7 a část BA 31 na vnitřní ploše hemisféry), multimodální, to znamená většímu množství smyslových systémů sloužící, asociační korová oblast, se při vybavování informací namáhá bez ohledu na informační obsah a smyslovou modalitu jejich kódování.
Vzdálenost v čase ovlivňuje při vybavování autobiografických vzpomínek činnost hipokampů. Čím vzpomínky starší, tím víc při jejich vybavování klesá aktivita pravého hipokampu, zatímco aktivita levého se nemění – zůstává vysoce aktivní při vybavování vzpomínek bez ohledu na jejich stáří. Dorzální části obou amygdal se při vybavování časově vzdálených vzpomínek chovaly podobně jako pravý hipokampus. Rozličná emoční intenzita a valence (kladná/záporná) vzpomínek na události v různých dobách přitom pokles aktivity amygdal úměrně stáří vzpomínek neměnila. To je pozoruhodné, protože amygdaly jsou klíčové uzly zpracování emočních informací jak co do intenzity, tak co do valence (8). To znamená kvalitu emoce obecně kladnou, nebo zápornou.
Z toho plyne v jak velké míře záleží na zadání úlohy. Mozek odpovídá pouze na to, na co se ho ptáme, a odpovídá na to přesně. I malé odchylky v zadání úlohy pochopitelně jeho činnost mění. Příkladem jsou odchylky činnosti mozku při testování autobiografické paměti fotografiemi snímanými jednou v zadaném prostředí univerzity, podruhé podobnými fotografiemi snímanými v laboratoři. Neuronální síť odpovídající epizodické paměti tvořená mediálními spánkovými a prefrontálními korovými oblastmi se aktivovala v obou případech. Fotografie z univerzitního prostředí však více než podobné fotografie z laboratoře aktivovaly mediální prefrontální kůru, jejíž činnost je jedním z podkladů sebeuvědomování, dále zrakovou a parahipokampální kůru odpovídající zrakové a prostorové paměti a také hipokampus, jenž se namáhá při vybavování vzpomínek (3).
Sémantická paměť
Sémantická paměť je druh deklarativní paměti, jejímiž obsahy jsou fakta, pojmy, významy slov i znalost předmětů. Sémantická paměť není závislá na kontextu, není tedy vázána na konkrétní čas a prostor. Jejím obsahům se musíme ze značné části vědomě učit, je sdílená kulturně.
Sémantický systém je tvořen rozsáhlými, do jisté míry vymezenými oblastmi levostranné týlní, temenní a čelní kůry na zevní i spodní ploše hemisféry. Tyto korové oblasti zpracovávají informace o tvaru i pohybu objektů, i o jejich užívání. Ve spánkovém laloku jsou v zadních částech zpracovávány generické informace, v čás-tech předních, včetně pólu, informace o unikátních objektech. Polární spánková kůra a jednotlivé korové oblasti levé dolní prefrontální kůry se podílejí na získávání, monitorování, výběru i uchovávání sémantických informací
Klinické zkušenosti dokazují, že mezi poruchami epizodické a sémantické paměti existuje dvojí disociace: například pacienti s Wernicke-Korsakovovým syndromem mají poškozenou epizodickou paměť, zatímco jejich paměť sémantická může být z valné části zachovaná. Na druhé straně existují pacienti s těžkým poškozením sémantické paměti, které dosahuje stupně označovaného jako sémantická demence, při poměrně zachované paměti epizodické. O tom, zda jsou všechny obsahy sémantické paměti nejprve součástí paměti epizodické a součástí sémantické paměti se stanou až po konsolidaci, nebo zda se jedná o dva, do jisté míry nezávislé druhy paměti, se diskutuje. Klinické zkušenosti však mluví spíše o druhé možnosti.
Amnézie
Amnézie je porucha deklarativní paměti, která se může týkat epizodické a sémantické paměti společně nebo zvláště. Kromě toho se může týkat událostí, které proběhly před dobou vzniku amnézie, to je retrográdní amnézie, ale i událostí, které se odehrají po době vzniku amnézie, to je anterográdní, také ortográdní nebo prográdní amnézie. Jiné druhy paměti při amnézii postiženy nebývají.
Amnestičtí pacienti jsou například schopni učení hybným dovednostem, mají tedy zachovanou procedurální paměť, aniž si pamatují, že se něčemu učili. Amnézie má řadu příčin a vzniká při poškození rozmanitých částí mozku.
Hipokampální amnézie
Klasickou kazuistikou je pacient H. M., kterému byly v r. 1953 oboustranně odňaty vnitřní části obou spánkových laloků počínaje temporálními póly v délce, jak nyní dokazuje magnetická rezonance, asi pěti centimetrů, včetně hipokampální formace, amygdaly a entorhinální kůry. Důvodem operace byla jinak neléčitelná epilepsie. Po operaci se četnost epileptických záchvatů snížila, ale:
„Tento mladý muž nebyl schopen poznat ošetřující personál, nenašel cestu do koupelny a zdálo se, že si z každodenních události svého života vůbec nic nepamatuje. Objevila se také částečná porucha paměti zasahující do minulosti. Nepamatoval si na smrt svého strýce, kterého měl rád a k níž došlo před třemi roky. Nepamatoval si vůbec nic z období, které prožil v nemocnici, i když byl schopen vybavit si několik běžných událostí, ke kterým došlo bezprostředně před jeho přijetím do nemocnice. Jeho rané vzpomínky byly velmi živé a nedotčené. Před deseti měsíci se jeho rodina přestěhovala do nového domu vzdáleného jen několik bloků od jejich starého domu a ještě k tomu ve stejné ulici. H. M. se dosud nenaučil novou adresu, i když si starou pamatuje dokonale. Cestu domů sám nenajde. Jeho matka říká, že H. M. den po dni luští stejnou křížovku, aniž by to mělo nějaký vliv na zlepšování výkonu. Čte znovu a znovu stejné časopisy, jejich obsah je pro něj stále nový…“ (11).
H. M. trpí globální anterográdní a částečnou retrográdní amnézií. Pracovní paměť postiženou nemá. Je schopen učení jednoduchým hybným dovednostem. Jeho stav se za celé desítky let, které od operace uplynuly, nezměnil. Rozsáhlá pokusná i klinická práce rozhodla teprve v následujících desetiletích, že kritickou strukturou, zúženým profilem informačního chodu, která rozhoduje o amnézii tohoto typu, je hipokampální formace.
- Následovali další pacienti, u nichž byla provedena klinicko-anatomická korelace, z níž plyne:
- Oboustranné poškození sektoru CA 1 hipokampu (obr. 2) dostačuje ke vzniku středně těžké anterográdní amnézie.
- Oboustranné poškození hipokampu, které postihuje i další hipokampální struktury, způsobí těžší anterográdní amnézii než poškození postihující jen sektor CA 1.

Poškození hipokampální formace může způsobit retrográdní amnézii zasahující období patnácti a více roků.
Nejčastější příčinou „hipokampální“ amnézie, se kterou se potkávají lékaři v každodennosti, je minimální kognitivní porucha (MCI) a vyvíjející se Alzheimerova nemoc. Tento typ poruchy recentní epizodické paměti se postupně prohlubuje do demence.
Diencefalická amnézie
Diencefalickou amnézii může způsobit jak poškození talamu, tak hypotalamu, případně jejich kombinace. Příčinou bývá Wernickeův-Korsakovův syndrom, vzácně trauma, v současnosti se tato amnézie často popisuje v souvislosti s ložiskovými ischemickými změnami. Korelátem amnézie u alkoholického Korsakovova syndromu je zmenšený objem hipokampu, corpora mamillaria a zvětšený objem III. mozkové komory. Naší zkušeností je, že nejčastější příčinou Wernickeova-Korsakovova syndromu je nedostatek thiaminu u lidí trpících malnutricí, nikoli závislost na alkoholu. Příčinou diencefalické amnézie obvykle doprovázené dalšími příznaky mohou být ložiskové cévní příhody. Amnézii při ložiskových cévních příhodách diencefala obvykle doprovázejí další příznaky. Kritickou strukturou je fasciculus mamillothalamicus propojující corpora mamillaria a nc. anterior thalami (obr. 3)
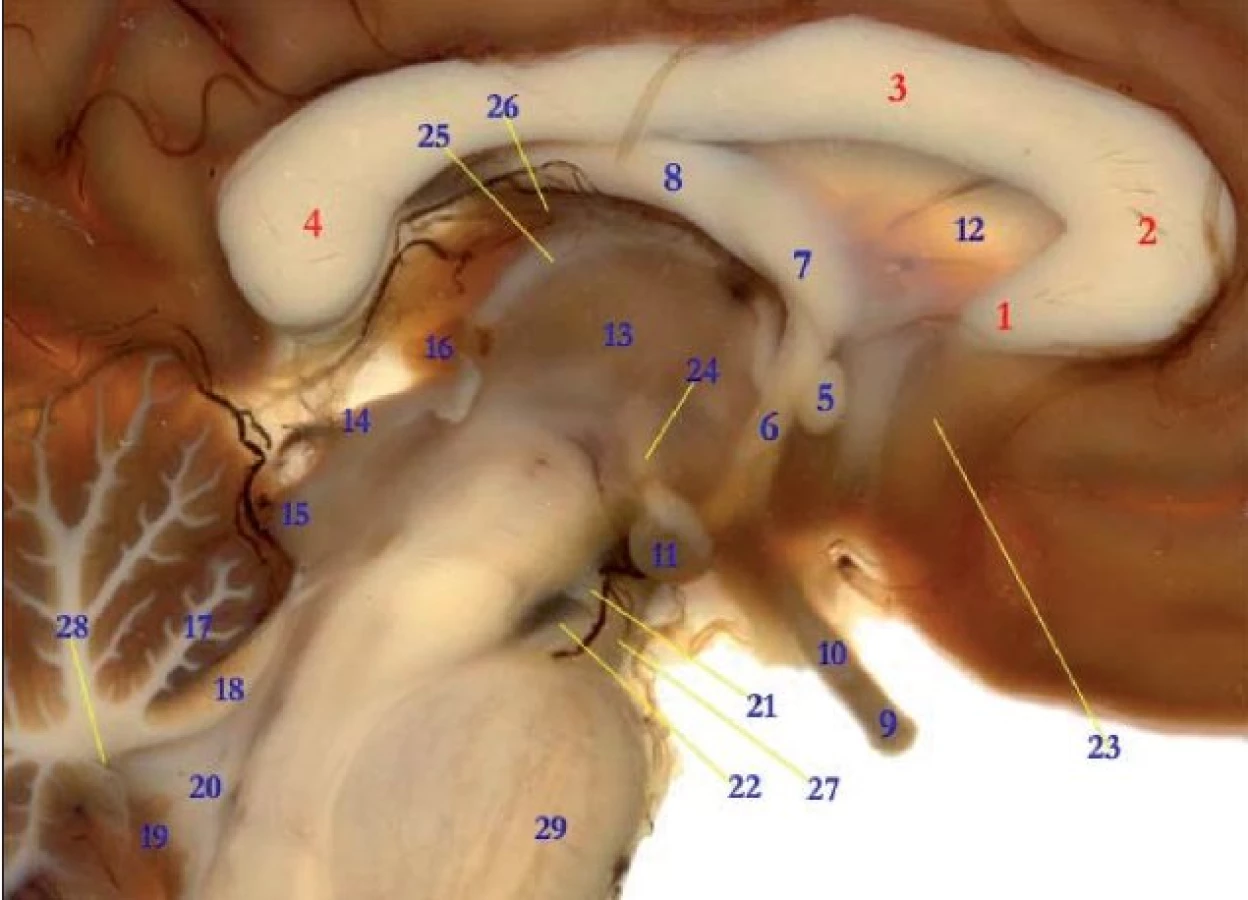
Amnézie jako důsledek poškození bazálního telencefala
Součástmi bazálního telencefala, oblasti při bázi mozku, která je „za a nad zkřížením zrakových nervů“, jsou laterální a mediální jádra septa u lidí mohutně vyvinutá, diagonální Brocův proužek, nc. basalis Menyerti, nc. accumbens (ventrální striatum).
Nejčastější příčinou ložiskového poškození této oblasti bývá prasklá výduť přední nebo spojovací mozkové tepny, případně následný nutný chirurgický výkon, vzácně úraz, zánět obvykle herpetického původu, primární a metastatické nádory. Větší poškození většinou zároveň postihne zadní orbitofrontální kůru, area subcallosa, případně přední části gyrus cinguli, přední části corpus callosum, a to souměrně, nebo nesouměrně. Charakteristicky je bazální telencefalon, zvláště cholinergní nc. basalis Meynerti, postiženo při Alzheimerově nemoci. Amnézie při poškození bazálního telencefala je podmíněna poškozením jeho cholinergních systémů propojených jak s hipokampem, tak s amygdalou a neokortexem.
Poškození orbitofrontální kůry, amygdaly, perirhinální kůry a mediálního hypotalamu, ať už v souvislosti s Wernickeovým-Korsakovovým syndromem, nebo z jiných příčin, mohou být spontánní konfabulace
Amnézie při ložiskovém poškození bílé hmoty
Ložiskové poškození soustav nervových vláken může způsobit amnézii v případech přerušení zúžených profilů informačního chodu diskonekcí (odpojením) funkčně významných soustav. Příkladem je amnézie, která může doprovázet porušení isthmus collateralis, svazku vláken, jehož zdrojem jsou vnitřní části spánkového laloku. Isthmus collateralis propojuje zadní části gyrus parahippocampalis s unimodálními a multimodál-ními smyslovými korovými oblastmi. Probíhá v hloubce sulcus collateralis. Přerušení tohoto svazku odpojuje informační vstup i výstup hipokampální formace. Příčinou bývají cévní mozkové příhody, nejčastěji různě velké infarkty, někdy i drobné, postihující oblasti zásobované a. cerebri posterior.
Amnézie se dále popisuje v souvislosti s poškozením korových a podkorových oblastí mozku v okolí zadní části (splenium) corpus callosum (obr. 3). Obvykle ji doprovázejí další příznaky. Místní poškození bývá opět nejčastěji důsledkem cévních mozkových příhod. Zda může způsobit amnézii i poškození fornixu, svazku vláken, jenž propojuje hipokampus zejména s hypotalamem, je předmětem probíhající diskuze. Chirurgické protětí fornixu případně spojené s poškozením přední komisury amnézii způsobit mohou.
Tranzitorní globální amnézie
Tranzitorní globální amnézie (TGA), která není vzácným onemocněním, se projevuje přechodným stavem v jehož průběhu si postižený jedinec není schopen zapamatovat nové informace. Jde tedy o anterográdní amnézii. TGA obvykle postihuje lidi, kteří jsou starší než padesát let, může se však objevit u lidí mladších. Porucha trvá jednu až osm hodin, nemá přesahovat 24 hodin. Pacienti jsou v jejím průběhu bdělí, komunikují, neztrácejí osobní identitu. Bývají zmatení, opakovaně se ptají, co se s nimi děje a kde jsou. Příčinou vzniku může být vazokonstrikce při hyperventilaci, tromboembolická ischémie i venózní kongesce postihující CA 1 oblast hipokampu (7).
Sémantická demence
Sémantická demence klinicky charakterizována selektivním poškozením sémantické paměti, které se projevuje těžkou anomií, poškozeným chápáním smyslu jednotlivých mluvených i psaných slov, poškozenou tvorbou příkladů v testech plynulosti sémantických kategorií („vyjmenujte co největší počet ptačích druhů…“), sníženou mírou obecných znalostí o lidech, předmětech a smyslu slov. Uchované jsou dalších složky chápání a tvorby jazyka, zejména fonologie a syntaxe, vizuopercepční a prostorové schopností a dovednosti, schopnost řešit neverbální úlohy jakož i autobiografická a epizodická paměť.
Příčinou sémantické demence jsou různé podoby frontotemporálních demencí, nejčastěji jde o frontotemporální demenci s ubikvitinovými inkluzemi (obr. 4) vzácnější příčinou jsou demence z komplexu Pickovy nemoci. Charakteristická je přitom levostranná převaha změn dokazující vazbu sémantického systému na levou hemisféru.

Priming
Priming (doslovně spěšná instrukce, podněcování, instruování předem) je anglický pojem pro druh nedeklarativní, nevědomé neboli implicitní paměti. Pokusné osobě se předvede nějaký podnět, například seznam slov. V časovém odstupu několika minut až několika hodin dostane zkoumaný jedinec trojhláskové kořeny těchto slov, které doprovází větší počet možných zakončení. Úlohou je doplnit příslušný kořen první koncovkou, která vyšetřovaného napadne. (Podnětem je například slovo motel, s odstupem času dostane vyšetřovaný jedinec seznam mo -týl, -ped, -tor…). Zdraví lidé zakončují „správnou“ koncovkou častěji než koncovkou „nesprávnou“. Didaktická klasifikace rozlišuje percepční a koncepční priming.
Percepční priming, neboli priming při vnímání, je co do smyslové modality specifický. Sémantické kódování jej neovlivňuje.
Koncepční priming, čili priming, jenž souvisí s tvorbou pojmů, modalitně specifický není, sémantické kódování jej ovlivňuje.
Ve skutečnosti bývá priming nějakým způsobem smíšený. Například v průběhu testování percepčního primingu si lidé často pomáhají sémantickým kódování podnětů. Vyšetřování funkčními metodami ukazuje, že jednoduchý percepční priming, jakým je doplňování kořenů slov, se projeví sníženou mírou aktivace korových oblastí, které se namáhaly v průběhu zpracovávání prvního podnětu. Priming lze chápat jako energetickou úsporu.
Pavlovovské podmiňování
U pokusných zvířat bylo zjištěno, že se pavlovovské podmiňování dá narušit poškozením nc. dentatus a nc. interpositus mozečku, mozečkové kůry, talamu, bazálních ganglií a gyrus cinguli. Poškození hipokampu pavlovovské podmiňování u zvířat i lidí nenarušuje, nicméně záznam činnosti jeho neuronů v průběhu podmiňování dokládá, že se jejich činnost mění, takže se procesu podmiňování účastní.
U lidí je při poškozeních mozečku pavlovovské podmiňování narušeno podobně jako u pokusných zvířat. Vyšetření zdravých dobrovolníků PET prokazuje, že se při pavlovovském podmiňování testovaném mrkacím reflexem aktivuje rozsáhlá neuronální síť:
- čelní kůra oboustranně,
- levý talamus,
- střední část pravé hipokampální formace,
- levý gyrus lingualis,
- pons, a
- oboustranně mozeček.
Pokles krevního průtoku byl zaznamenán oboustranně ve spánkové kůře, v pravém putamen a v zevním částech hipokampální formace. S postupem učení vzrůstal krevní průtok v nucleus caudatus, hipokampální formaci, gyrus fusiformis levé hemisféry, v pravé spánkové kůře, ve Varolově mostu a v mozečku. Charakteristické je, že s pokračujícím učením, a tedy vyšší mírou automatického zpracovávání informace, klesal krevní průtok v prefrontální kůře Jinak řečeno: kůra se úměrně tomu, jak učením zvládla úlohu , „méně namáhá“.
Psychogenní amnézie a syndrom falešné paměti
Obvykle se rozlišují čtyři druhy psychogenní amnézie:
- Disociativní amnézie, kterou charakterizuje neschopnost vybavit si významné, obvykle traumatizující nebo stresující osobní informace, přičemž porucha je natolik rozsáhlá, že ji nelze vysvětlit běžnou zapomnětlivostí.
- Disociativní fuga, kterou charakterizuje náhlé a nečekané opuštění domova nebo místa, kam postižený obvykle chodí do zaměstnání, doprovázené neschopností vybavit si vlastní minulost, jakož i poruchou pocitu osobní identity, často i domněle novou identitou.
- Disociativní porucha identity (dissociative identity disorder), dříve nazvanou mnohočetná porucha osobnosti (multiple personality disorder), pro niž je typická přítomnost dvou nebo většího počtu odlišných identit nebo stavů osobnosti, které opakovaně přebírají kontrolu nad chováním postiženého jedince. Stav doprovází neschopnost vybavit si osobně významné informace, která je natolik rozsáhlá, že ji nelze vysvětlit běžnou zapomnětlivostí.
- Depersonalizační porucha, při které mají postižení lidé opakovaný pocit, že jsou odloučeni od vlastních duševních procesů nebo od vlastního těla, jejich schopnost ověřovat skutečnost je však nedotčená.
Syndrom falešné paměti (false memory syndrome) se týká „vybavení“ živých vzpomínek na události, které se nestaly. Jde o výrazně medializovaný syndrom, jenž se objevuje u dospělých lidí, kteří obviňují své rodiče z údajného sexuálního zneužívání. To mělo proběhnout v dětství. Na zneužívání tito lidé „zapomněli“. Údajně si je vybavili v průběhu psychoterapie, kterou začali navštěvovat pro potíže odlišného druhu. V 90tých letech minulého století se zejména v USA, méně v Británii, objevila „epidemie“ těchto poruch, která posléze odezněla.
Činnost mozkové kůry při vybavování iluzorních vzpomínek
Vzpomínky nejsou otiskovou reprezentací proběhlých událostí. Vznikají tím, že se složí z informací, které jsou rozmístěny v mnoha vzdálených místech mozku. Jejich vznik je závislý na řadě minulých i současných vlivů. Desítky let je známo, jak často a snadno se vzpomínky zkreslí. Vybavování informací z paměti je rekonstruktivní proces, čemuž nasvědčují i výsledky zkoumání činnosti mozkové kůry funkčními zobrazovacími metodami.
Vznik iluzorních vzpomínek na jevy, které se nestaly nebo na informace, s nimiž se vyšetřovaní zdraví jedinci nesetkali, se dá snadno uměle navodit, například pokusem, v němž se užije seznam slov a slovních „návnad“.
Činnost mozku při rozpoznávání skutečných a iluzorních vzpomínek byla ověřována řadou experimentů. V jednom z nich bylo úkolem dobrovolníků rozlišovat slova, která se naučili, od slov, která byla blízká, ale neučili se jim, a nových slov, která k oběma předešlým skupinám neměla vztah. Zjistilo se, že levý hipokampus aktivují jak skutečné, tak falešné položky, předpokládá se, že jeho činnost souvisí s vybavováním informací bez ohledu na to, zda jsou skutečné nebo falešné. Naproti tomu zadní část levého parahipokampálního závitu aktivovaly víc skutečné než falešné položky. Podobně odlišně se aktivovaly oblasti prefrontální kůry: dorsolaterální prefrontální kůru aktivovaly oboustranně skutečné a falešné položky více než položky nové – předpokládá se monitorování vybavované informace.
Levou dorsolaterální prefrontální kůru aktivovaly víc nové položky než položky skutečné nebo falešné, předpokládá se podíl na zpracování sémantické informace. Precuneus a zevní temenní korové oblasti více aktivovaly pravé a falešné než nové položky. Naproti tomu orbitofrontální kůru a mozeček aktivovaly více falešné než skutečné položky. Z toho plyne, že přední úseky mediálního spánkového laloku mezi skutečnými a falešnými položkami nerozlišují, zatímco zadní části mezi nimi rozlišují (2).
MUDr. František Koukolík, DrSc.
Oddělení patologie a molekulární
medicíny
Národní referenční laboratoř
prionových chorob
Fakultní Thomayerova nemocnice s poliklinikou
Vídeňská 800
140 59 Praha 4 Krč
E-mail: frantisek.koukolik@ftn.cz
Sources
1. Baddeley, A. The fractionation of working memory. Proc. Natl. Acad. Sci., 1996, 93, p. 13468-13472.
2. Cabeza, R., Rao, S.M., Wagner, A.D. et al. Can medial temporal lobe regions distinguish true form false? An event-related functionl MRI study of veridical and illusory recognition memory. Proc. Natl. Acad. Sci. USA 2001, 98, p. 4805-4810.
3. Cabeza, R., Prince, S., Daselaar, S. M. et al. Brain activity during episodic retrieval of autobiographical and laboratory events: an fMRI study using novel photo paradigm. J. Cognitive Neuroscience 2004, 16, p.1583-1594.
4. Gadian, D.G., Alcardi, J., Watkins, K.E. et al. Developmental amnesia associated with early hypoxic-ischaemic injury. Brain 2000, 123, p. 499-507.
5. Hort, J., Rusina, R. a kol. Paměť a její poruchy. Paměť z hlediska neurovědního a klinického. Praha: Maxdorf Jessenius, 2007. ISBN 978-80-7345-004-5.
6 Koukolík, F. Funkční systémy lidského mozku. Norma a poruchy. 3. přepracované a rozšířené vydání. Praha: Galén, 2008 (v tisku).
7. Lee H.Y., Kim, J.H., Weon, Y-Ch. et al. Diffusion-weighted imaging in transient global amnesia exposes the CA1 region of the hippocampus. Neuroradiology 2007, 49, p. 481-487.
8. Maguire, A. E., Frith, Ch.D. Lateral asymmetry in the hippocampal response to the remoteness of autobiographical memories. J .Neurosci. 2003, 23, p. 5302-5307.
9. Milner, B., Squire, L.R., Kandel, E.R. Cognitive neuroscience and the study of memory. Neuron, 1998, 20, p. 445-468.
10. Scoville, W.B., Milner, B. Loss of recent memory after bilateral hippocampal lesions. J. Neurol. Neurosurg. Psychiatry 1957, 20, p. 11–21.
11. Summerfield Ch., Greene, M., Wager, T. et al. Neocortical connectivity during episodic memory formation. PloS Biology 2006, vol.5. e128 DOI:10.1371/journal.pbio.0040128 (www.plos-bio-logy.org).
12 Willingham, D.B. Systems of memory in the human brain. Neuron 1997, 18, p. 5-8.
Labels
General practitioner for children and adolescents General practitioner for adultsArticle was published in
General Practitioner

2008 Issue 2
Most read in this issue
- Memory and its disturbances
- Current possibilities and problems of antiretroviral therapy
- The hoarding of possessions and animals
- Acute toxic hepatitis following consumption of a herbal preparation